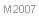|
|||||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||
|
Género
Cataglyphis
©Kiko Gómez & Xavier Espadaler |
Género muy diverso dentro de la región mediterránea que se caracteriza por los palpos maxilares larguísimos y que sobrepasan claramente el borde inferior de la mandíbula, así como por el espiráculo propodeal en forma de ranura. En los machos, se da una gran variedad en la genitalia masculina, carácter empleado por Agosti (1990) para la reordenación del género y que es de gran ayuda en la identificación específica.
Las características que definen al género, siguiendo Agosti (1990) son para las reinas y obreras:
En los machos:
En el mismo documento, Agosti abandona la división en subgéneros y propone otra basada en nueve grupos de especies, de los que cuatro están presentes en la Península Ibérica. Sus características principales en las obreras son (ver definición completa y características de los machos en Agosti 1990): Grupo albicans: En las obreras, peciolo nodiforme con la cara dorsal anterior plana y anguloso en vista lateral; en las reinas tiene forma de escama. El gastro elevado a 90º en locomoción. Márgenes del propodeo con quetas erectas. Tercer segmento del palpo maxilar comprimido, y el cuarto segmento individualmente de la misma longitud o menor que el quinto y el sexto juntos. MPI (longitud 4º segmento maxilar/longitud 5 º segmento maxilar*100)>90. Poco polimórficas, con las obreras mayores de longitud menor que 3,2 mm. Normalmente uniformes en la coloración, de negro brillante a amarillento brillante. Si son coloreadas, el gastro es más oscuro que el resto del cuerpo. Grupo altisquamis: Peciolo en forma de cuña. Color uniformemente negro, uniformemente castaño-amarillento o bicoloreadas con el gastro negro y el resto del cuerpo de negro a rojo claro. Mates. Escultura de la cabeza finamente reticulada, a veces con puntuación en la frente. Tercer segmento maxilar engrosado en su parte distal. MPI<80. Tamaño muy grande en las majors, con longitudes de mesosoma (ML) de hasta 5 mm. Grupo cursor: Peciolo en forma de escama, con las caras anterior y posterior formando un ángulo pero formando una cresta solo de manera parcial. Negras, de apariencia mate. Excepcionalmente brillantes y/o con una tonalidad castaño-rojiza en la cabeza. La cabeza presenta una estriación fina en sentido loginudinal. En caso de ser reticulada, el dorso del mesosoma no presenta pilosidad. Tercer segmento maxilar redondeado. Tercer y cuarto segmentos maxilares individualmente de longitud menor que el quinto y el sexto juntos. MPI<90. majors con longitud del mesosoma menor de 3,4 mm. Grupo emmae: Peciolo en forma de escama, con las caras anterior y posterior formando un ángulo pero formando una cresta solo de manera parcial. Negras a castaño oscuras, mates. Mandíbulas con un diente apical y y otro preapical seguidos de dos o tres dentículos, de forma que a veces solo presenta cuatro dientes. Dimórficas, con obreras de ML=2 mm y soldados de ML=2,8 mm. tercer segmento maxilar de sección redondeada. Tercer y cuarto segmentos maxilares individualmente menores que el quinto y el sexto juntos. Esta división en grupos no es, ni mucho menos estanca, sino que, como muchas otras veces, intentan ser "cajas" donde ir agrupando toda la diversidad existente en la naturaleza. Un ejemplo claro en Cataglyphis es C. floricola, que si bien es atribuida a este grupo por el Dr. Tinaut (1993), basandose en características de la genitalia del macho, no comparte en la casta obrera características como el color oscuro, la dentición, la longitud relativa de los palpos maxilares o la ausencia de dimorfismo. Resumiendo, que estos grupos de especies, si bien pueden llegar a ser útiles, no son, ni mucho menos, estancos.
Gran parte de este género ha sido reordenado en profundidad por el Dr. Alberto Tinaut (1990a, 1990b, 1990c) para todos los grupos excepto para el grupo albicans, que ha tenido diversas aportaciones recientes por parte de los Dres. De Haro y Collingwood (1995 -con P. Douwes-, 1996, 2000, 2001, 2003), si bien este grupo todavía adolece de una revisión integral para la totalidad de la fauna ibérica, y podría seguir aportando sorpresas en el futuro.
Las decisiones que hemos tomado respecto de ciertas citas dudosas o con necesidad de clarificación han sido las siguientes:
A nivel mundial se conocen más de 70 especies. Son eminentemente paleárticas, desde Portugal a Mongolia y el Ganges y desde el Sur de Sudán a Eslovaquia. Prefieren las estepas y hábitats mediterráneos, evitando los bosques. No son especies especialistas en vivir en los desiertos, aunque aparecen a menudo en los bordes de los mismos y en los oasis. Presente desde los 0 a los 3.000 metros de altitud. En la Península ibérica la mayor diversidad de especies se da en el Sur de la Península Ibérica, si bien C. Ibericus amplía su distribución hacia lugares más bien cálidos del Norte de la Península Ibérica y C. piliscapus está distribuida por el Sur de Francia/Norte de la Península. En cuanto a su origen, todas las especies son endémicas, excepto la citada C. piliscapus.
Las especies bombycinus, lucasi y kurdistanicus son dimórficas. No se conocen obreras para C. hannae, inquilina de C. bicolor Estas hormigas viven en las zonas templadas y cálidas (estepas, páramos, desiertos...) y claramente termófilas. C. fortis, por ejemplo, es una especialista en recoger los cadáveres de los insectos muertos por el calor. Hay casos donde forrajean a tan elevada temperatura que la comida está disponible sólo para ellas. Los nidos se ubican bajo tierra, aprovechando así las condiciones aislantes del terreno. En la especie C. bicolor se han medido temperaturas exteriores del terreno de 70 ºC, mientras que en el interior del nido la temperatura a 22 cm de profundidad era de entre 30 y 35 ºC, mientras que a 40 cm. Era prácticamente constante a 30 ºC. Además emplean tácticas de enfriamiento como recubrir las entradas de sus nidos con guijarros, vegetación, trozos de ramas, etc, de modo que capturan el calor de sol aislando al nido. Este comportamiento daría base a las afirmaciones de Plinio y Herodoto de las hormigas mineras de oro de las cercanías de Kabul, al recoger los guijarros de los aluviones de los ríos de la zona, muy ricos en pepitas de oro. Son eminentemente carroñeras (con excepciones), con unos patrones bien definidos de comportamiento. Forrajean practicamente a lo largo de todo el día, si bien concentran su actividad a lo largo de 2-3 horas, cerrando el nido durante la noche. Se ha demostrado que emplean la memoria visual para guiarse, tomando referencias tanto del terreno como de la posición del sol "leyendo" la luz polarizada, llegando a alejarse hasta 100 metros del nido. Se han descrito dos especies parásitas, una de C. bicolor: C. hannae, que carece de obreras y habita en el desierto del Sahara y otra de C. stipes: C. zakharovi en Turkmenistan.
En la Península Ibérica la guía imprescindible son los trabajos del Dr. Tinaut (1990a, 1990b, 1990c) para todos los grupos excepto para el grupo albicans. Este grupo, si bien todavía en estudio, tiene como referencias los trabajos de los Drs. De Haro y Collingwood (1995 -con P. Douwes-, 1996, 2000, 2001, 2003). La definición dentro de la tribu Formicini, así como su estructura en grupos de especies ha sido estudiadas por Donat Agosti (1990, 1994) y es la guia actual para la ordenación de este género. Si analizamos la fauna paleártica y mediterránea, existen trabajos de revisión en Emery (1906), que ya ofrece las primeras claves a machos basadas en genitalia, complementada por él mismo en 1908 con la revisión del grupo viaticus. Estos trabajos continúan con el Dr. Santschi (1929), que ofrece la primera división en subgéneros basándose entre otros, en la forma de los palpos maxilares, caracter empleado en todavía en la actualidad para separar los grupos de especies. Tras estos trabajos, no se vuelve a realizar un trabajo que revise el grupo en su conjunto, restringiéndose la mayoría bien a determinadas zonas geográficas (Petrov 1986; Radchenko 1997, 1998) o a determinados grupos de especies (Wehner, 1981; Wehner et al. 1994). Existen claves actualizadas en internet de la región sub-sahariana en Ants of West Africa. Agosti, D. (1990). Review and reclassification of Cataglyphis. Journal of Natural History, 24: 1457-1505 Agosti, D. (1994). The phylogeny of the ant tribe Formicini (Hymenoptera: Formicidae) with the description of a new genus. Systematic Entomology, 19: 93-117 De Haro, A.; Collingwood, C. A., Douwes, P. (1995). Nota preliminar sobre sistemática molecular gen-aloenzimática de algunas formas españolas y marroquíes del grupo albicans del género Cataglyphis (Hym., Formicidae). Orsis (Organismes i Sistemes), 10: 73-81 De Haro, A.; Collingwood, C. A. (1996). Estudio comparado de la genitalia de dos poblaciones españolas de Cataglyphis (Hym., Formicidae), grupo albicans de color negro. Orsis, 11: 53-58 De Haro, A.; Collingwood, C. A. (2000). Cataglyphis douwesi sp. nov. Del grupo albicans de color negro de Cádiz, en la costa suratlántica de la Península Ibérica. Orsis, 15: 57-67 De Haro, A.; Collingwood, C. A. (2001). Cataglyphis sp. Nueva especie del grupo albicans de color negro del Cabo de Gata (Almería), España. Orsis, 16: 89-92 De Haro, A.; Collingwood, C. A. (2003). Cataglyphis gadeai sp. nov. (Hym. Formicidae), del grupo albicans de color negro del Cabo de Gata (Almería), SE de España. Orsis, 18: 19-27 Emery, C. (1906). Rassegna critica delle specie paleartiche del genere Myrmecocystus. Memorie della Reale Accademia delle Scienze dell'Istituto di Bologna, (6)3: 47-61 [pagination of separate: 173-187] Emery, C. (1908). Myrmecocystus viaticus et formes voisines. Bulletin de la Société Vaudoise des Sciences Naturelles, 44: 213-217 Petrov, I. Z. (1986). Distribution of species of the genus Cataglyphis Foerster, 1850 (Formicidae, Hymenoptera) in Yugoslavia. Arh. Biol. Nauka, 38: 11-12 Radchenko, A. G. (1997). A review of ants of the genus Cataglyphis Foerster (Hymenoptera, Formicidae) from Asia. Entomological Review (Washington), 77 (6): 684-698 Radchenko, A. G. (1998). A key to ants of the genus Cataglyphis Foerster (Hymenoptera, Formicidae) from Asia. Entomological Review (Birm.), 78 (4): 475-480 Tinaut, A.; Plaza, J. L. (1990a ("1989")). Situación taxonómica del género Cataglyphis Förster, 1850 en la Península Ibérica. I. Las especies del subgénero Cataglyphis Förster. EOS (Revista española de entomología), 65(1): 189-199 Tinaut, A. (1990b). Taxonomic situation of the genus Cataglyphis Förster, 1850 in the Iberian Península. II. New position for C. Viatica (Fabricius, 1787) and redescription of C. Velox Santschi 1929 stat. n.. EOS (Revista española de entomología), 66(1): 49-59 Tinaut, A. (1990c). Situación taxonómica del género Cataglyphis Förster, 1850 en la Península Ibérica. III. El grupo de C. Velox Santschi, 1929 y descripción de C. humeya sp. n.. EOS (Revista española de entomología), 66(2): 215-227 Tinaut, A. (1993). Cataglyphis floricola nov. sp. new species for the genus Cataglyphis Förster, 1850 (Hymenoptera, Formicidae) in the Iberian Peninsula. Mitteilungen der Schweirzerischen Entomologischen Gesellschaft, 66: 123-134 Wehner, R. (1981). Verhaltensphysiologische, funktionsmorphologische und taxonomische Studien an zwei Arten der Cataglyphis albicans-Gruppe (Formicidae, Hymenoptera). Mitteilungun der Deutschen Gesellschaft fur Allgemeine und Angewandte Entomologie, 3: 214-217 Wehner, R.; Wehner, S.; Agosti, D. (1994). Patterns of biogeographic distribution within the bicolor species group of the North African desert ant, Cataglyphis Foerster 1850. Senckenbergiana biologica, 74: 163-191
|
||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|